

Fisiología de cultivos
Photosynthesis performance and leaf water potential impairments of Tahiti Lime affected by Wood pocket
Desempeño de la fotosíntesis y disminución en el potencial hídrico de las hojas de Lima Tahití afectadas por Wood pocket
 Eleonora Rodriguez lrodriguezp@agrosavia.co
Jairo García L
Javier O Ordu
Eleonora Rodriguez lrodriguezp@agrosavia.co
Jairo García L
Javier O Ordu
Temas Agrarios
Universidad de Córdoba, Colombia
ISSN: 0122-7610
ISSN-e: 2389-9182
Periodicity: Semestral
vol. 25, no. 2, 2020
Received: 01 September 2020
Accepted: 27 December 2020

Abstract: The progressive death of Tahiti lime trees in productive stage was occasioned for la physiopaty Wood pocket (WP) observed in the production region of north-central Tolima (Colombia) in 2012, which led to significant detriment in fruit production of 79.98% with respect to the year 2011. The presence and level of damage caused by WP has been associated with dry warm climate conditions related mainly to high temperatures, a similar condition that occurred in this producing area. Little is known about the causes of decreased foliar gas exchange and leaf water potential in plants affected by WP; hence, this study aimed to investigate the relationships between physiological parameters (leaf water potential and foliar gas exchange) of Tahiti lima plants with symptoms of WP, and hydrodynamic and humidity conditions of soil in the area. The evaluation was performed in a commercial plantation of Lima Tahiti located in the municipality of Flandes. The values in photosynthetic parameters and leaf water potential, even in healthy plants, were lower than those reported for this species and the presence of WP decreases in greater proportion. The results indicated that the hydrodynamic properties, low availability of water in the soil and weather conditions contribute to the development of WP. We hypothesized that the constant water deficit and high temperatures cause rupture of tracheids and cavitation in the xylem, deteriorating the vascular system causing the death of wood or WP.
Keywords: A/Ci curves, Gas exchange, Physiological behavior, Abiotic disorders, Physiopathy, Inter-Andean valleys, A, Ci curves, Ci curves.
Resumen: La muerte progresiva de los árboles de lima Tahiti en etapa productiva fue ocasionada por la fisiopatía Wood pocket (WP) observada en la región productora del centro-norte del Tolima (Colombia) en 2012, lo que provocó un detrimento significativo en la producción de frutos del 79,98% en relación con el año 2011. La presencia y nivel de daño causado por WP se ha asociado a condiciones de clima cálido seco relacionado principalmente con altas temperaturas, condición similar a la que ocurre en esta zona productora. Se sabe poco sobre las causas de la disminución del intercambio gaseoso y el potencial hídrico de las hojas en plantas afectadas por WP, por lo tanto, este estudio tuvo como objetivo investigar las relaciones entre los parámetros fisiológicos (potencial hídrico foliar e intercambio de gases foliar) de las plantas de lima de Tahití con síntomas de WP con las condiciones hidrodinámicas y humedad del suelo en la región de cultivo de Lima. El efecto de WP y las condiciones hidrodinámicas y humedad del suelo, sobre los parámetros fotosintéticos y el potencial hídrico foliar se determinó en un ensayo experimental en una plantación comercial de Lima Tahití ubicada en el municipio de Flandes. Los valores en parámetros fotosintéticos y potencial hídrico foliar incluso en plantas sanas fueron inferiores a los reportados para esta especie y la presencia de WP los disminuyo en mayor proporción. Nuestros resultados indicaron que las propiedades hidrodinámicas, la baja disponibilidad de agua en este suelo y las condiciones climáticas son favorables para el desarrollo de WP. Planteamos la hipótesis de que el constante déficit hídrico y las altas temperaturas provocan la rotura de las traqueidas y la cavitación en el xilema deteriorando el sistema vascular provocando la muerte de la madera o WP.
Palabras clave: Curva A/Ci, Intercambio gaseoso, Comportamiento fisiológico, Desorden abiótico, Fisiopatía, Valles interandinos.
Colombia produces an average of 79.200 ton of Tahiti lime (Citrus latifolia Tanaka) per year of which the Santander Department is the highest producer with 62.706 ton, followed by Antioquia, Tolima and Risaralda with productions of 9.223 ton, 4.236 ton and 2297 ton, respectively. The highest yield ha-1 observed in Santander was 21.16 ton (Agronet, 2018). The cultivation of Lima Tahiti in Colombia is mainly for fresh consumption and processed products, for local and international markets because there are no sanitary restrictions for its exports (DANE, 2015).
In 2011, Tolima department reported 2.853 ha harvested Tahiti lime, with a production of 60.508 ton and an average yield of 28 ton ha-1. These values decreased dramatically in 2012, when only 820 ha were harvested, with 12.300 ton fruit production and an average yield of 15 ton ha-1, showing a 79.68% decline in production. This decrease in production was attributed to the presence of Wood Pocket (WP), a physiological disorder or physiopathy caused by genetic and abiotic factors such as hifg temperatures causing sectorized death in Tahiti, also called Persian or Bearss trees, leading to their total extermination (ICA, 2011).
WP is a worldwide serious problem that is not generally recognized (Calavan, 1957). Its presence has been reported in the United States, Mexico, Belize, Saudi Arabia, Oman, Egypt and Colombia (Knorr, 1957; Almeida et al., 2000; ICA, 2011). Its presence in Mexico, the world's leading producer of Tahiti and Persians, showed the severity of the damage by reducing the profitability of the fruit and processed products (Almeida et al., 2000).
Symptoms of WP include leaf yellowing with characteristic leaf blotch, striking fruit chimeras, dieback and cracking in trunks and branches. The main evidence in the wood is the stain that could be observed when a section is cut in a branch or trunk. If all of these symptoms are present in a declining lime tree, then there is a presence of the WP disorder (Roistacher, 2000). The life expectancy of lime trees is proportionally inverse to the prevailing high temperature. For example, in Saudi Arabia or Oman, where weather is constantly hot, lime trees affected by this physiopathy die within 2 to 4 years. In Mexico, trees need to be replanted within 5 to 10 years while in Belize they live for 8 -10 years. In United States, bears lime trees are replaced within 8 to 15 years. WP symptoms are more severe and develop faster under warm temperatures, being particularly drastic in desert regions (Roistacher, 2000).
The effect of WP disorder may be devastating, as exemplified by the destruction of the Florida Tahiti lime industry in the 1950´s (Fawcet & Calavan, 1947). Fortunately, through the selection of two resistant materials of Tahiti lime with good fruit characteristics and absence of WP symptoms, the Tahiti lime industry in Florida revived. These selections have developed symptoms of WP in different regions of the world that experience elevated temperatures, reason why its widespread use is limited.
In Mexico, WP disorder was reported in 2003 in the citrus growing region of Martínez de la Torre, Veracruz. Currently, WP is present in all states that produce this citrus species (Veracruz, San Luis Potosí, Tabasco, Yucatán, Colima, Jalisco, Nayarit, and others). This abnormality progressively reduces tree vigour and affects fruit quality because produces localized spots. WP has been described in Persian lime and Lisbon lemons (Citrus limon). There are no agronomical practices for limiting WP damage in citrus trees and micro-grafting does not eliminate further development of symptoms. However, evidence from recent research suggests that damage may be reduced by rigorous selection of native genotypes tolerant to WP (INIFAP, 2013).
In Colombia, wood pocket was reported in the region where Lima Tahiti is produced, located at the center of the Tolima department, causing death of trees once they reach their productive stage (ICA, 2011). This region presents characteristics of dry weather condition with high average temperatures, high level of evaporation and crop evapotranspiration (ETc), as well as high variation in the daily temperature delta and high value of accumulated heat units (AHU) (Hernández et al., 2014).The region of the inter-Andean valley of the municipality of Espinal (Tolima, Colombia), corresponds to a tropical dry forest (TDF) life zone, with bimodal rainfall regime with an accumulated annual precipitation of 1,476 mm (IGAC, 2004). The first rainy season occurs between March and May, also the months with highest precipitation (202 mm). The second rainy period occurs between October and November, with October having the highest rainfall of 176 mm. The dry season occurs also in two cycles, being the first between December and February where January is the driest. The second dry season is between June and September, of which August is the month with the highest temperature (Hernández et al., 2014).
Little is known about the causes of decreased foliar gas exchange and leaf water potential in plants affected by WP, hence, this study aimed to investigate the relationships between physiological parameters (leaf water potential and foliar gas exchange) of Tahiti lime plants with symptoms of WP with hydrodynamic conditions and humidity of soil in the Lima growing region of the department of Tolima, Colombia.
MATERIAL AND METHODS
Study site
One field experiment was conducted in a farm named Yulima in the municipality of Flandes in Tolima department, Colombia, planted with 4-year old Tahiti lime trees, located at 04°14´06.38" N, 075°53.054´ W, 315 m above sea level, with average temperature of 33 ± 5 °C and relative humidity of 40 ± 5 %. The experiment was performed from August to October, 2013. The chemical characteristics of the Inceptisols of the experimental area were as follows: pH (H2O) 5.93; P, 133 mg dm–3; K, 0.52 cmol dm–3; Ca2+, 6.78 cmol dm–3; Mg2+, 1.1 cmol dm–3 and Al3+, 0.0 cmol dm–3. Concentration of Mo in the soil was 2.75 mg kg-1 of soil. Apparent high density (1.626 -1.769) and sandy loam soil.
The orchard was maintained using conventional practices performed locally for Lima Tahiti crops, including fertilization before sowing based on results of the soil chemical analysis. The soil was fertilized with nitrogen, K2O, Ca, Mg, SO4, Mn and Zn at 276, 120, 44,18.8, 6.8, 6.4 and 6 kg ha-1, respectively. The nutritional requirements were applied based on the nutrient balance index methodology considering the recommendations presented by Corrales (2002) for soil optimal nutrient concentration, and the recommendation of Sánches et al. (1994) for optimal nutrient concentration in leaf tissue.
Sprinkler irrigation was applied at a volume of 36 l of water hour-1 for a period of three hours per day, which corresponded to a total volume of 108 l day-1, with an application frequency of three times week-1, this irrigation volume corresponds to the traditional amount of water applied to the crop in the region.
Weather variables were measured using a meteorological station Hobo U12-006 (Coltein Ltda, Bogotá, Colombia), installed in the plot coupled with a data logger decagon em50 for measuring volumetric content of water and soil hydraulic conductivity, precipitation, relative humidity and average temperature.
Hydrodynamic characteristics of the soil
Soil moisture and retreating water were established by the method of McQueen and Miller (1968). The rate of soil infiltration was determined using the ring-infiltrator method (FAO, 2006). The volumetric soil moisture content was monitored using a datalogger decagon em50. Penetration resistance was measured using an Agridry Rimik penetrometer, model CP 20, following the methodology reported by ASAE (1992).
Leaf gas exchange
Net carbon assimilation rate (A), stomatal conductance to water vapor (gs), internal-to ambient CO2 concentration ratio (Ci/Ca) and transpiration rate (E), were measured in fully expanded leaves with an infrared CO2/H2O gas analyzer (LICor 6400, Lincoln, NE) equipped with a blue/red light source (Li-6400-02B). Actual Water Use Efficiency (WUE) was calculated as A/E and actual intrinsic water use efficiency was calculated as A/gs (Machado et al., 1994). Measurements were conducted at environment temperature (27 ± 0.9°C) and CO2 conditions, with artificial light (1000 μmol photons m−2 s−1), from approximately 08:00 to 12:00 h at 0, 30 and 60 days of observation (dao), (Bispo et al., 2016 a). Measurements were performed in six leaves from a group of 10 plants of each treatment, selected from those that had the most visible symptoms of the disease at each evaluation time
Daily record measurements of Leaf gas exchange and Leaf water potential
All of the above measurements were taken in ranges of one hour, between 7:00 and 17:00 for two days at 0, 30 and 60 dao.
Experimental design
Two independent experiments were carried out in a complete randomized design, with two treatments (plants without symptoms, PWS, and plants with 25% of damage by wood pocket, PWP). Each experiment was performed using ten plants per treatment (60 in total), in each sampling time (0, 30 and 60 dao). Each experimental unit consisted of one Tahiti lime tree of 4 years old. Data from A, gs, Ci/Ca, E, A/E, A/gs and Ψp registered in the two experiments were combined after determination of homogeneity of variances by Cochran’s test. Plant disorder severity was assessed monthly by calculating the percentage of necrotic tissue respect to the total area of the tree.
Statistics
All data recorded were subjected to analysis of variance (ANOVA). For each sampling time, ten plants from each treatment were assessed for Ψp and gas exchange measurements. Means were compared by the T test (P ≤ 0.05) using SAS statistical package (Release 8.02 Level 02 M0 for Windows, SAS Institute, Inc., 1989, Cary, NC, USA).
RESULTS AND DISCUSSION
Hydrodynamic characteristics of the soil
The characterization study of the hydrodynamic properties of soils suggested limitations in the movement and retention of water at Yulima farm. Physical infiltration, moisture retention, and apparent soil density indicated an average infiltration rate of 15.98 mm hour-1, that is classified as moderately slow to rapid (Table 1). They were also characterized for capacity to store available water between low and very low at the four depths at which samples were taken (Table 2). Hydraulic conductivity was classified as slow indicating resistance to water flow through soil profile (Table 3).



Mechanical resistance to soil penetration in the Yulima farm presented values between 4 and 5 Mpa, showing physical limitation from 15 cm depth in the soil profile, with high restriction to roots development (Pla, 1998), therefore inducing shallow roots, which can cause overturn and limitations of plants to abosrb water and nutrients favoring WP (Whiteley and Dexter, 1982). In dry soil conditions, such as those observed in Yulima farm, the development of a more densely rooted layer at 30 cm depth soil, or less, is conducive to maintaining the water supply in the plant (Yu et al., 2007).
Values of field capacity (FC) and permanent wilting point (PWPO) were 26% and 22.5%, respectively; the value of 26% in CC indicates low water retention and 22.5 % in the PMP indicates low moisture available in the soil. The volumetric soil moisture content indicates a constant water deficit with values lower than PMPO. Cumulative precipitation in the five months of observation was 439.80 mm (Figure 1).
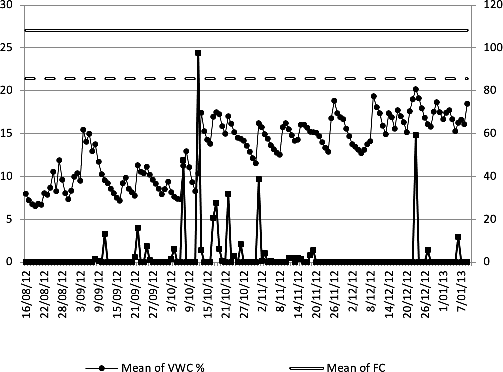
The study of hydrodynamic properties of the soil indicated limitations in the movement and retention of water to a value of usable water in the soil of 3.5% (Figure 1). Application of 324 l water week-1 plus an accumulated precipitation of 489.2 mm were insufficient for an appropriate water supply, since the volumetric content of water in the soil (VWC) remained below the point of permanent wilting (PWPO) during the conduction of the experiment (Figure 1). Considering the low moisture retention capacity and the moderately slow basic infiltration present in the soil at Yulima farm and as one of the main factors inducing WP is defficiency in available water, the recommended irrigation methods would be pressure irrigation using techniques such as dripping or micro-sprinkling which eventually allow the supply of small irrigation sheets with high frequencies. Accordingly, the effects of root size and architecture on final yield will depend on the distribution of soil moisture and the level of competition for water resources within the plant community (King et al., 2009).
Gas exchange
To the extent of our knowledge, this is the first study that estimates the severity of WP related to alterations in water relations and photosynthesis of Tahiti lime trees. Characterization of the hydrodynamic properties and soil moisture content, along with environmental factors such as temperature, relative humidity and precipitation, indicated that during the period of the study, a constant water deficit may induce constant stress in lime trees even in PWS, although PWP presented lower values of A, gs, E, Ci/Ca, A/E, A/gsand Ψp.
| Parameters | Sources of variation PWS -PWP | |
| 0 dao 30 dao | 60 dao | |
| A | * ns | * |
| gs | ** ns | * |
| Ci/Ca | * ns | ns |
| E | ** ns | * |
| A/E | ns * | ns |
| A/gs | ** ns | ns |
| ns: not significant. |

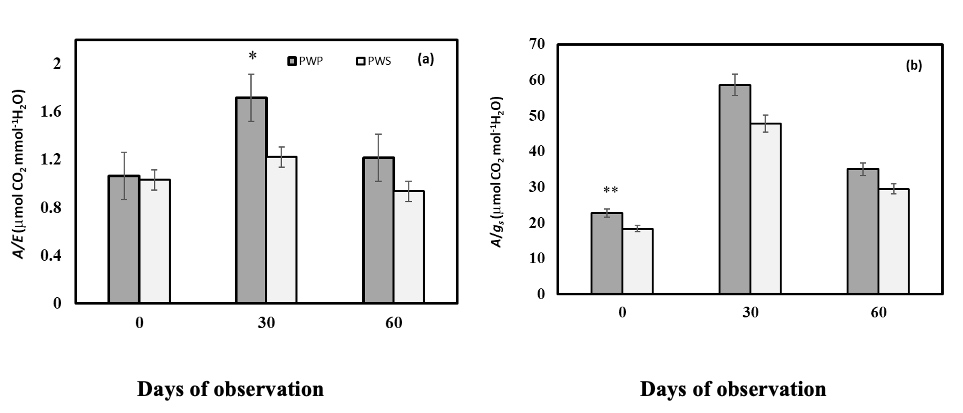
Significant differences (P ≤ 0.05) between PWS and PWP for A, gs, Ci/Ca and E, were observed for all sampling times (0, 30 and 60 dao) (Table 4). PWP presented average decreases of 19% for A, 34% for gs, 25% for E and a 5% for Ci/Ca (considering the data from 0, 30 and 60 dao), compared to PWS (Fig. 2a, b, c, d). Overall, PWP showed higher A/gs and A/E ratios (Fig. 3a, b).
The main visible effect of water stress in lime trees is growth decrease of the aerial part, reduction of foliar area and increase of root growth. Growth decrease of the aerial part is caused by stomata closing, which is regulated by the plant hormone abscisic acid (ABA), (Mcdowell, 2011; Jdey et al. 2014). When stomata are closed and water loss by transpiration is reduced, the CO2 input decreases reducing photosynthesis and consequently, increases stomatal resistance (1/gs) (Azcón and Talón, 2008; McDowell et al., 2008). Plant growth is finally diminished by decreasing of turgor pressure, which limits leaf expansion and proliferation of new shoots (Allen et al., 2010; Skirycz and Inzé, 2010; Anderegg, 2015).
Leaf water potential
PWP presented an average decrease of 17%, based on 0, 30 and 60 dao data, compared to PWS (Figure 4a). Overall, PWP showed higher Ψp during the day (Fig. 4b). Tahiti lime PWS and PWP values of Ψp are very high (Fig. 5a and 5b), which indicates that the plants are making a great effort elaborating solutes and osmolytes that do not allow them to lose water (osmotic adjustment) (Fig. 5b). Osmotic Adjustment (OA) is a metabolic process entailing a net increase in intercellular solutes in response to water stress (Morgan, 1984; Zhang et al., 1999; Serraj and Sinclair, 2002). As soil moisture declines, OA favors turgor maintenance, and hence the integrity of metabolic functions (Tuberosa, 2012).
Overall PWP showed higher values of Ψp during the day (Fig. 5b). A measure based on plants, such as water potential (ψ), should be the most direct indicator of water stress of the plant and, therefore, of the need for irrigation (Nio et al., 2011).
Daily record measurement of Leaf gas exchange
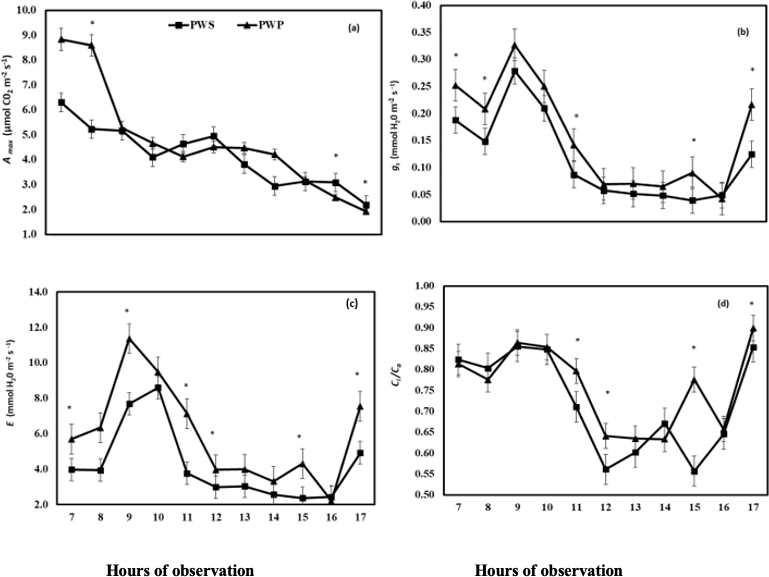
Wood pocket led to devastating effects on A. Wood pocket was able to limit the impairment of photosynthetic gas exchange as indicated by the highest values of A in PWS during the day (Fig. 5a). The values of gs, E and Ci/Ca were significantly higher at 11, 12, 15 and 17 hours, respectively, for PWP in comparison to PWS (Fig. 5b, c and d). For PWS and diseased PWP plants, the transpiration behavior (E) was similar, with highest values at early morning (Fig. 5c). Our results indicated that when water deficit conditions, the presence of Wood pocket contributes to a higher reduction of A, gs, E, Ci/ Ca (Fig. 2 and 5).
In relation to gas exchange in healthy plants (PWS) and diseased plants (PWP), it was observed that rate of carbon liquid assimilation (A) is very low in both, when compared to other perennial fruit trees, presenting values lower than 9 μmol CO2 m-2 s-1 (Whiley et al., 1999), with a decreasing pattern throughout the day, and without the normal reduction due to the increase of solar radiation in the morning hours (Fig. 5a). These values are lower than those reported under optimal environmental conditions (Syvertsen and Lloyd, 1994) which normally varies between 12 and 14.5 μmol m-2 s-1 (Medina, 2003). This difference was probably caused by water deficit conditions in the soil together with variability in environmental elements such as air temperature and relative humidity at levels lower than optimal physiology (Allen et al., 2010; Hoffmann et al., 2011;Cochard and Delzon, 2013).
Values of Ci/Ca ratio have a similar behavior in PWP and PWS (Fig. 5d). Photosynthesis is a critical process for crop growth and performance, and it can be dramatically impaired when leaves are infected by pathogens (Berger et al., 2007) or physiopathies. A decrease of the net carbon assimilation rate in infected leaves has been attributed to decreases in mesophyll and / or stomatal conductance of CO2, impairments in the rates of biochemical reactions of photosynthesis, changes in the number or structure of chloroplasts or changes in the secretion of phytotoxic chemicals into asymptomatic tissue (Berger et al., 2007).

According to accumulated historical records of climatic variables between 1981 and 2010, the landscape of the north-central Tolima region is characterized by having a negative water balance (precipitation vs evapotranspiration) from six to nine months a year, usually from November to March and from June to September (Hernández et al., 2014). In addition, the soil in this area is highly drained because formation from materials from volcanic activity of Machín mountain and the fluvial dynamics of Magdalena River, and its tributaries (Cortolima, 2006).
The genetic material of Tahiti lima planted in Colombia more probably was originated from a single clonal accession (Rodriguez et al., 2017). The presence of WP, only in the producing of north-central Tolima region, maybe is associated with management conditions of plantations, especially the hydric and edaphoclimatic conditions (Hernández et al., 2014), rather than the propagation material. All mentioned conditions lead to a region environmentally predisposed to WP development (Ríos et al., 2018). In fact, this was illustrated with the drastic decline of harvest between 2016 (2853 ha) and 2011 (420 ha) (Agronet, 2018).
According to the nutritional diagnosis carried out during 2018 in 20 Tahiti lime farms in the north-central Tolima region, similar results in foliar and soil nutrient levels were found. For both parameters i) deficiencies in Mg, Zn and N, ii) excess in B, Fe, P, Mn, and Cu and, iii) optimal condition in S, Ca and K were observed (Rodríguez et al., 2018). The low amount of nutritional deficiencies is due to the frequency and quantity increase of fertilizers applied to crops as a management strategy against WP by farmers. The negative effect of inadequate edaphoclimatic conditions, on plant growth and water availability, stimulates the magnitude of water deficit, which can severely affect Tahiti lime crops in Tolima department (Pérez-Pérez et al., 2010; Vélez et al., 2012). Management strategies recently developed by Ríos et al. (2020) ratified the decrease in damage caused by WP due to the use of 150% nutritional requirement from soil and leaf applied fortnightly. This fertilization strategy and keeping moisture content in soil closest to the field capacity improves the functional and physiological response of the PWS and PWP trees. It is been associated with the increase of gs and A which in turn, rise fruit yield and renew foliage during the crop season.
CONCLUSIONS
The results indicated that weather conditions, hydrodynamic properties, and low availability of water in the soil favored the development of WP. Further research on irrigation systems, the application of antiperspirant products, mitigation factors such as moisture retainers and, the addition of organic matter are necessary to improve moisture retention in the soil profile. In addition, native and imported genotypes tolerance to water deficit should also be considered during future assessments.
Acknowledgments
ACKNOWLEDGMENTS
To the Ministry of Agriculture and Rural Development for financing this study and to Mr. Alfonso Ricón, owner of the Yulima farm for his collaboration for the establishment and development of research on his property.
REFERENCES
Agronet. 2018. Red de Información y comunicación estratégica del sector agropecuario – AGRONET Colombia. Ministerio de Agricultura y Desarrollo Rural. https://www.agronet.gov.co/estadistica
Almeida, I.H., Iracheta, M.M., Jasso, J., Curti, S.A., Ruíz, P., and Rocha, M.A. 2002. Reexamination of citrus viroids of Tahiti lime in Mexico. Revista Mexicana de Fitopatología 20(2):152-160.
Allen, C.D., Macalady, A.K., Chenchouni, H., Bachelet, D., and Mcdowell, N.A. 2010. Global overview of drought and heat- induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 259:660-684. Doi: 10.1016/j.foreco.2009.09.001
Anderegg, W.R.L. 2015. Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation. New Phytol 205:1008–1014. Doi: 10.1111/nph.12907
ASAE, Standards. 1992. S 313.4. Soil cone penetrometer. St. Joseph. 611p.
Azcón, J., and Talon, M. 2008. Fundamentos de fisiología vegetal, segunda/Ed. Mc Graw Hill/Interamericana de España.
Berger, S., Sinha, A.K., and Roitsch, T. 2007. Plant physiology meets phytopathology: plant primary metabolism and plant- pathogen interactions. Journal of Experimental Botany, 58(15–16): 4019–
Bispo, W. M. S., Araujo, L., Moreira, W. R., Silva, L.D. and Rodrigues, F. A. 2016a. Differential leaf gas exchange performance of mango cultivars infected by different isolates of Ceratocystis fimbriata. Scientia Agricola, 73 (2): 150- 158. Doi: 10.1590/0103-9016-2015-0022
Calavan, E.C. 1957. Wood Pocket disease of lemons and seedless limes. Citrograph 42: 265-268, 300-304.
Cochard, H., and Delzon, S. 2013. Hydraulic failure and repair are not routine in trees. Annals of Forest Science 70:659-661. Doi: 10.1007/s13595-013-0317-5
Corporación Autónoma Regional del Tolima (Cortolima). 2006. Plan de ordenación y manejo de la cuenca mayor del río Coello. https://www.cortolima.gov.co/estudios
Corrales, A., 2002. Manual ilustrado para la producción de cítricos en Colombia. Asocítricos. Ministerio de Agricultura y Desarrollo Rural. Convenio MADR- Asocítricos 167 pp
Departamento Administrativo Nacional de Estadística (DANE). 2015. Boletín mensual INSUMOS Y FACTORES ASOCIADOS A LA PRODUCCIÓN AGROPECUARIA
FAO. 2006. Evapotranspiración del cultivo. Guías para la determinación de los requerimientos de agua de los cultivos. Bol. (56) Roma
Fawcet, HS., and Calavan, E.C. 1947. Wood pocket, a newly reported disease of lemons. Phytopathology 37:843.
Hernández, D.R., Mateus, D., and Orduz, J.O. 2014. Características climáticas y balance hídrico de la lima ácida Tahití (Citrus latifolia Tanaka) en cinco localidades productoras de Colombia. Revista Colombiana de Ciencias Hortícolas, 8(2): 217-229.
Hoffmann, W.A., Marchin, R.M., Abit, P., and Lau, O.L. 2011. Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought. Glob Change Biol 17:2731-2742. Doi: 10.1111/j.1365-2486.2011.02401.x
ICA, 2011. Muestreo fitosanitario de los cítricos en el Tolima. Informe de actividades. Semestre B de 2011. Gerencia seccional Tolima, Área de protección vegetal.
INIFAP, 2013. Simposio internacional sobre HLB en cítricos ácidos. http::///D:/Descargas/4126%20 Memoria%20Simposio%20HLB%20 en%20citricos%20acidos%20(1).pdf
Instituto Geográfico Agustín Codazzi (IGAC). 2004. Estudio general de suelos y zonificación de tierras departamento de Tolima [Memoria técnica]. Bogotá, Colombia.
Jdey, A., Slama, I., Rouached, A., and Abdelly, C. 2014. Growth, Na+, K+, osmolyte accumulation and lipid membrane peroxidation of two provenances of Cakile maritima during water deficit stress and subsequent recovery. Flora 209, 54–62. Doi: 10.1016/j.flora.2013.10.002
King, C. A., Purcell, L. C., and Brye, K. R. 2009. Differential wilting among soybean genotypes in response to water deficit. Crop Sci. 49, 290–298
Machado, E.C., Quaggio, J.Á., Lagoa, A.M., Ticelli, M., and Furlani, P.R. 1994. Trocas gasosas e relacaoes hídricas em laranjeiras com clorose variegada dos citros. Rev. Bras. Fisiol. Veg. 6, 53–57.
Mcdowell, N.G., Pockman, W.T., and Allen, C.D. 2008. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytologist 178:719-739.
Mcqueen, I.S., and Miller, R.F. 1968. Calibration and evaluation of a wide- range gravimetric method for measuring moisture stress. Soil Science, 106 (3): 225-231.
Medina, CL. 2003. Fisiologia da produção. In: Mattos JR, De Negri JD, Figueiredo JO. (Ed.). Lima ácida Tahiti. Campinas: Instituto Agronômico. p. 67-80.
Morgan, J. W. 1984. Osmoregulation and water stress in higher plants. Annu. Rev. Plant Physiol. 35: 299–319.
Nio, SA., Cawthray, G.R., Wade, L.J., and Colmer, T.D. 2011. Pattern of solutes accumulated during leaf osmotic adjustment as related to duration of water deficit for wheat at the reproductive stage. Plant Physiol Biochem 49: 1126–1137.
Ríos, L., Correa J.F., Rojas, C.A., and Dorado, D.Y. 2018. Caracterización edafoclimática de la zona productora de lima ácida Tahití (Citrus latifolia Tanaka) en el Tolima (Colombia), afectada por una fisiopatía. Ciencia y Tecnología Agropecuaria,19 (3):545-567.
Ríos, L., Chaali, N., Jaramillo, C.I., Ouazaa, S., Correa, J.F. 2020. Irrigation and nutrition as criteria for adequate management of Tahiti acid lime trees affected by a physiological disorder in tropical conditions. Scientia Horticulturae, 270- 109438. Doi 10.1016/j.scienta.2020.109438
Rodriguez, D., Murcia, N., and Martinez, M. 2017. Variabilidad genética de Limas ácidas con marcadores Microsatélites Amplificados al Azar (RAMs) en Colombia. En Memoria. V simposio Internacional de Fruticultura tropical y subtropical IX simposio internacional de Piña. FRUTICULTURA.
Rodríguez, E., Gutierrez, J.S., Orduz, J.O. 2018. Diagnostico nutricional del cultivo de la lima ácida Tahití [Citrus latifolia (Yu Tanaka) Tanaka] en el departamento del Tolima (Colombia). Temas agrarios, 23 (2): 144-153.
Roistacher, C. 2000. Wood Pocket - A genetic disorder. Citrograph 45 :(3):3-6.
Sánches, A., VanRaij, B., Blasco, E., Malavolta, E., Vitti, G., Cantarella, H., Quaggio, J., Teofilo, J., De Negri, J., Rodríguez, O., Bataglia, O., 1994. Recomendaçoes de adubação e calagem para citros no Estado de São Paulo. rev. e atualizada. Laranja (Cordeirópolis) ed. especial. 27 p.
Skirycz, A., and Inzé, D. 2010. More from less: plant growth under limited water. Current Opinion in Biotechnology 21: 197-203.
Syvertsen, J.P., and Lloyd, J. 1994. Citrus. In: Schaffer B, Andersen PC (eds), Handbook of Environmental Physiology of Fruit Crops: Sub-Tropical and Tropical Crops, pp.65-99. CRC Press, Boca Raton.
Tuberosa, R. 2012. Phenotyping for drought tolerance of crops in the genomics era. Front Physiol 3:347, 1-26.
Whiley, A.W., Searle, C., Schaffer, B., and Wolstenholme, B.N. 1999. Cool orchard temperatures or growing trees in containers can inhibit leaf gas exchange of avocado and mango. Journal of the American Society for Horticultural Science 124:46-51.
Whiteley, G.M., and Dexter, A.R. 1982. Root development and growth of oilseed, wheat and pea crops on tilled and non- tilled soil. Soil & Tillage Research, 2:379– 393.
Yu, G., Zhuang, J., Nakayama, K., and Jin, Y. 2007. Root Water Uptake and Profile Soil Water as Affected by Vertical Root Distribution. Plant Ecology, 189 (1)15-30. http://www.jstor.org/stable/40212846
Zhang, J. X., Nguyen, H. T., and Blum, A. 1999. Genetic analysis of osmotic adjustment in crop plants. J. Exp. Bot. 50:291–302.
Additional information
Conflict of Interest: The authors declare that
it is an original work and there was no conflict of interest of any kind in the elaboration and publication
of the manuscript.

